ТРИПАНОСОМЫ
Спасибо нашим инвесторам из казино онлайн
ТРИПАНОСОМЫ
, ТРИПАНОСОМИАЗ. Трипаносомы, паразитические простейшие из рода Trypanosoma Gruby, 1843, сем. Trypanosomidae, порядка Protomonadina, класса Mastigophora. К семейству Trypanosomidae относятся близкие роду Trypanosoma роды Leptomonas, Phyto-monas, Leishmania, Herpetomonas, Crithidia и Schizotrypanum (см. рисунок). Последний род многими включается в род Trypanosoma. Трипаносомы паразитируют в крови и в тканях млекопитающих (включая человека), птиц, пресмыкающихся, земноводных и рыб. В крови трипаносомы представляются в виде весьма подвижных образовании продолговатой формы. Лишь
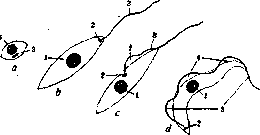
Схема морфологических типов семейства Trypanosomidae: а— лейшмания; Ь—лептомонада (Lepto-monas); с—критидия; d—трипаг. осома; -/—ядро; 2— блефаробласт; г—жгут; d— волнообразная перепонка.
нек-рые виды, как Т. rotatorium лягушек, на определенных стадиях развития имеют форму плоских образований, ширина к-рых немного уступает длине. Наиболее мелкие трипаносомы имеют в длину от 12—15
мм
до 25—30
мм
(средний размер ряда патогенных трипаносом человека и др. млекопитающих). Более крупные формы, как напр. непатогенная трипаносома крупного рогатого скота, Т. theileri, достигают 60—70
мм.
Встречаются и трипаносомы, достигающие в длину 120—130—200
мм
(трипаносомы оленей, рыб). Тело трипаносом одето тонкой оболочкой, пелликулой (перипласт). Пе-рипласт обладает азурофилией и в препаратах, энергично окрашенных по Романовскому, принимает красноватый оттенок, маскирующий голубой оттенок базофильной плазмы. У большинства трипаносом перипласт не обнаруживает особых структур, но у некоторых, напр. у трипаносомы амфибий, обладает продольной исчерченностью. По одному краю вдоль тела трипаносомы перипласт приподнят жгутом, начинающимся у заднего конца трипаносомы и б. ч. заходящим за передний конец ее тела. Приподнятая жгутом над собственно плазмой дупликатура перипласта образует т. н. волнообразную перепонку или ундулирующую мембрану. У основания жгута, у заднего конца трипаносомы, сидит т. н. базальное тельце, часто сливающееся с
более
крупным зерном, т. н. парабазальным телом. Парабазалыюе толо (а иногда совокупность его с базальиым зерном) называется блефаробласт, кинетонуклеус, ки-петопласт. Блефаробласт красится, как ядро, и дает положительную реакцию Фейльгена. .Ядро расположено центрально, оно имеет кариосому и периферически расположенный хроматин. Некоторые трипаносомы в естественном состоянии не имеют парабазального тела (Т. equinum). У других в норме небольшой процент экземпляров оказывается лишенным его. В условиях эксперимента удается получить безблефаро-бластиые расы трипаносом путем воздействия красок (пиронин и др.). На разных стадиях инфекции и в культурах в плазме трипаносом обнаруживаются мелкие зерна округлой формы азурофильной природы, близкие к волютину. В организме переносчиков и зараженных животных трипаносомы могут менять свою морфологию, принимая формы, свойственные близким родам Leishmania, Leptomorias, Crithidia. Наиболее близко к собственно трипаносомной форме подходит форма критидии, отличающаяся от трипаносомы тем, что у нее блефаробласт расположен впереди ядра, соответственно чему ундулирующая мембрана оказывается более короткой (см. рисунок). Удлиненные формы со свободным жгутом, отходящим от блефаробласта, расположенного у передней вершины клетки, лишенные волнообразной перепонки, обозначаются как лептомонадные формы (Leptomonas, см. рисунок). Округлые или овальные мелкие формы с ядром и блефаробластом, но без жгутика, обозначаются как лейшманиевидные формы (см. рисунок). Размножение трипаносом происходит путем деления. Половой процесс для трипаносом не установлен. Делению ядра обычно предшествует деление блефаробласта, причем жгут остается за одной из половин базального зерна с примыкающей половиной парабазального тела, а от второй половинки блефаробласта начинает отрастать новый жгут, идущий параллельно старому и скоро достигающий той же длины. Деление ядра происходит митотическим путем. За ним следует продольное расщепление тела. В некоторых случаях наблюдается ряд повторных делений ядра и блефаробласта с образованием множественных особей. В разные периоды инфекции размножение трипаносом протекает с разной интенсивностью. В периоды оживленного размножения трипаносом в крови животного встречаются экземпляры более длинные, со свободным жгутом, и более широкие и короткие формы, у которых жгут заканчивается у переднего конца тела. В другие периоды встречаются гл. обр. формы однородные по своей морфологии. Ряд работ был посвящен диференциации разных видов трипаносом по их длине. Позднейшие исследования показали, что эта длина меняется в зависимости от фазы инфекции и вида животного, которому привита данная трипаносома. В силу значительного морфол. сходства ряда патогенных трипаносом многих видов животных и чрезвычайно выраженной способности трипаносом к изменению своих биол. свойств в зависимости от длительности пассирования на одном или разных видах животных, разграничение отдельных видов патогенных трипаносом в ряде случаев представляет большие трудности. Лаверан и Мениль (Laveran, Mesnil) ввели метод диференцировки трипаносом путем т. н. перекрестного заражения, основанный на том, что животные, перенесшие заражение определенным видом трипаносом и спонтанно выздорове
вшие, оказываются иммунными к данному виду, но восприимчивыми к другим видам трипаносом. Ими же введен и серологический метод идентификации трипаносом, основанный на том, что сыворотка иммунного к данному виду трипаносом животного предохраняет от заражения гомологичным видом трипаносом, но не действительна в отношении гетерологич-ных видов. На практике однако оба метода представляют ряд затруднений в виду того, что в пределах каждого вида трипаносом имеются многочисленные штаммы, иммуыологически отличные друг от друга, а также в связи с тем, что. иммунитет после ликвидации инфекции держится недолго. Диференциация разных видов трипаносом иногда возможна на основании их разного отношения к хим.-терап. веществам и специфичности переносчика—беспозвоночного хозяина. Первые попытки культивирования трипаносом принадлежат Данилевскому(1886— 1889), наблюдавшему размножение трипаносом птиц и лягушек в капилярах с кровью. Настоящие культуры трипаносом с пересевами впервые были получены Нови и Мек Нил ем (Novy, Mac Neal) на среде из мясопептонного питательного агара, к к-рому при t° в 50° добавлялось от 10 % до 200—300% дефибринированной крови крысы, собаки или кролика. Рост происходил преимущественно в конденсационной жидкости при t° 25—37°, В культурах морфология трипаносом упрощается: они дают формы крйтидий и лептомонад. Последние могут образовывать розетки, совершенно сходные с розетками в культурах Leishmania. Культуры Т. lewisi сохраняли свою вирулентность для крыс после ряда пассажей в пробирках при комнатной tc в течение года и более. Те же авторы получили также культуру Т. brucei, а вслед за ними Лаверан и Мениль вырастили Т. evansi. Первоначально предложенная Нови и Мек Нилем среда была упрощена Никол ем (см.
Лейшманиозы).
На этой среде NNN удается выращивать ряд трипаносом (Т. theileri, Schizotrypanum cruzi и др.). Т. theileri, T. melophagium и близкие к ним культивируются и на бульоне с кровью. Нек-рые виды трипаносом нуждаются в добавлении к среде глюкозы. Неллер (Noller) культивирует трипаносом на поверхности мясопептонного агара с 10% глюкозы, смешанного пополам с дефибринированной кровью. На поверхности такого агара трипаносомы птиц, Т. theileri и др. дают весьма характерные формы роста в виде колоний со своеобразными боковыми выростами. В культурах при комнатной t° трипаносомы дают формы, аналогичные стадиям их развития в беспозвоночном хозяине. При повышении t° до 37° они принимают форму, свойственную им в организме теплокровного. Культивирование патогенных форм трипаносом удается лишь с большим трудом, причем трипаносомы быстро теряют свою вирулентность. Беренс (Behrens) культивировал Т. brucei на агаре, приготовленном на наваре из гороха. Отдельным авторам удавалось получать культуры Т. congolense, Т. brucei и Т. gambiense [см.
Сонная болезнь
и отдельную таблицу (т. XXXI, ст. 855—8j6, рис. 5)] на среде Понселя—дефи-бринированная инактивированная кровь кролика с добавлением дест. воды и на других модификациях его среды, содержащих разные количества соли, пептон и желатину. Культивирование трипаносом нередко применяется как метод обогащения для обнаружения трипаносом при весьма малом их содержании в периферической крови. Неллер в& 1916—28 гг. применил этот метод для установления переносчика Т. theileri: получив культуры трипаносом из задней кишки слепней, он заразил этими культурами телят. Пути р а с п р о странен и я трипаносом специфичны для каждого вида. Известны следующие механизмы передачи трипаносом: 1) контактный путь—непосредственная передача от животного к животному при контакте слизистых оболочек; 2) контаминативныйпуть— через поедание животным промежуточного хозяина или испражнений последнего, содержащих вирулентные стадии трипаносом; 3) ино-кулятивный путь—через укус промежуточного хозяина, вводящего в кровь животного заразные стадии трипаносом. Промежуточное положение между вторым и третьим способом составляет заражение через внедрение в ранку, произведенную укусом, патогенных стадиев трипаносом, попавших на кожу животного по соседству с ранкой вместе с испражнениями насекомого. Контактным путем передается Т. equi-perdum (см.
Дурина).
Контаминативным путем распространяются трипаносомы группы lewisi (Т. melophagium, lewisi и др.). Переносчиком для Т. melophagium является овечий рунец (Ме-lophagus ovinus), для Т. lewisi—крысиная блоха (Ceratophyllus fasciatus). В средней кишке насекомого трипаносомы подвергаются делению и затем превращаются в критидиальные формы. Последние в прямой кишке насекомого после ряда повторных делений снова принимают форму трипаносом. Эти т. н. метациклические т
рипаносомы выходят наружу вместе с экскрементами насекомого и заражают животное, поедающее насекомое целиком или слизывающее с шерсти его экскременты. Т. о. для трипаносом с контаминативным способом распространения характерно развитие заразных стадиев в заднем отделе кишечника насекомого. При заражении инокулятивным способом заразные стадии трипаносом локализуются в конечном счете в переднем отделе пищеварительного тракта (в хоботке) насекомого или пиявки. Для разных видов трипаносом с инокулятивным способом передачи характерны разные типы локализации и развития паразитов в насекомом. Для одних видов трипаносом насекомое является лишь механическим переносчиком, так напр. Т. equinum передается через разные виды слепней, переносящих на хоботке заразную кровь с больного животного на здоровое. Другие виды трипаносом (Т. congolense, gambiense) проделывают сложный цикл развития в передних отделах пищеварительного тракта и в средней кишке насекомого (Т. rotatorium в пиявке), заканчивающийся появлением метациклических трипаносом, внедряющихся у одних видов в слюнные железы, а у других возвращающихся в хоботок, минуя слюнные железы. В этих случаях от момента сосания заразной крови до момента, когда насекомое становится заразным, должен пройти нек-рый промежуток времени, б. ч. в несколько дней, к-рый необходим для того, чтобы трипаносомы завершили свой цикл развития в промежуточном хозяине. Б-ни, вызываемые трипаносомами, обозначаются как трипаносомоз ы (менее правильно—как трипаносомиазы или трипанозы). Практический интерес представляют преимущественно трипаносомозы человека и домашних животных. Трипаносомозы человека рас-■ пространены только в тропических странах:
сонная болезнь
(см.)—-в Африке,
Шагаса болезнь
(см.)—в Америке. Трипаносомозы животных имеют значительно более широкое распространение, но по преимуществу», в силу условий распространения, особенно свирепствуют в тро – пических, субтропических и степных областях, где нередко причиняют столь большой вред животноводству, что делают совершенно невозможным разведение и содержание скота в ряде областей Африки и Индии. Важнейшие трипаносомозы животных.
Дурила (см.).
—С у – а у р у, трипаносомоз верблюдов. В СССР распространен в Среднеазиатских республиках, в Казакстане, уральских и астраханских степях. Возбудитель Т. ninae kohl-yakimov. Б-нь может иметь острое, подострое и хроп. течение. Острые формы ведут к гибели животного в полтора-два месяца. Характерные симптомы—вялость, конъюнктивит, отеки. Подострые и хроп. формы ведут к сильному истощению, к концу б-ни развиваются парезы, паралич зада. Аналогичное заболевание, вызываемое тем же возбудителем, встречается в уральских степях и у лошадей.—С у р р а, заболевание, сильно распространенное в Индии и в Сев. Африке, спорадически заносимое в Америку, Австралию, Южную Африку и др. страны. Поражает преимущественно однокопытных, Заболевают также верблюды, слоны, собаки, у к-рых б-нь имеет б. ч. смертельный исход. Возбудитель Т. ovansi. Основными переносчиками являются двукрылые из родов Та-banus, Haematopota и Stomoxys. В Зап. Африке аналогичное заболевание известно под названием Mbori.—М у р р и н а, заболевание лошадей и мулов Центр. Америки. Возбудитель Т. hippicum, сходен с Т. evansi. Передается механически через мух.—М aide Caderas, заболевание лошадей и ослов в тропической зоне Юж. Америки. Сопровождается гематурией. Возбудитель Т. equinum отличается отсутствием парабазалыгого тела.—Н а г а н а или нгана (син. Baleri, Aino), распространенное в Центр. Африке смертельное заболевание лошадей, ослов, собак, поражающее и других домашних животных, у к-рых имеет однако более легкое течение. Возбудитель Т. brucei цере-дается через муху цеце из рода Glossina; очень близок к возбудителям трипаносомоза человека—Т. gambiense и Т. rhodesiense. Патогенез и симптоматология трипаносомозов. Течение и патогенез разных трипаносомозов представляют очень много общего. При остром течении инфекции трипаносо-мы во многих случаях безудержно размножаются в крови животного до самой его смерти. Смерть животного наступает в короткий срок: 2 недели—2—3 мес. При хрон. течении нередко удается установить вначале период массового размножения трипаносом в крови с резко выраженным лихорадочным состоянием, опуханием лимф, желез, отеками. За ним следует длительный период, в течение к-рого трипано-сомы то появляются в периферической крови (рецидивы) то исчезают на длительный срок. В этом периоде также наблюдаются подъемы t°, развивается анемия, общее истощение, скопление жидкости в полостях тела. В да
льнейшем появляются тяжелые изменения со стороны нервной системы, параличи и псих, расстройство. Смерть наступает в состоянии кахексии. В патогенезе заболевания помимо токсических продуктов трипаносом известную роль повиди-мому играет и развивающаяся при усиленном размножении трипаносом гипогликемия как результат потребления трипаносомами углеводов крови. У многих видов животных вслед за острым периодом или с самого начала инфекция принимает весьма легкое течение. Живот – | ные, кровь к-рых остается заразной, что можно доказать перевивкой значительных количеств крови на свежих животных, практически здоровы и трудоспособны. Вместо с тем такие животные оказываются устойчивыми против реинфек-ции тем же штаммом (нестерильный иммунитет, premunition). В других случаях может наступить полное выздоровление животного, которое после полной ликвидации паразитов сохраняет на более или менее продолжительный срок иммунитет к гомологичным трипаносомам.— Один и тот же вид трипаносом вызывает у разных видов животных неодинаковую картину болезни. Острые формы трипаносомозов чаще | наблюдаются у однокопытных и верблюдов; у I рогатого скота—чаще подострые формы и хронические с доброкачественным течением. Патологическая анатомия. Являясь по преимуществу паразитами крови, три-паносомы поражают также определенные ткани и системы органов. Значительное развитие трипаносом имеет место в лимф, путях и железах, в к-рых они иногда развиваются в огромном количестве еще до появления в крови. Нек-рые трипаносомы образуют значительные скопления в разных органах: трипаносомы лягушек в почках, трипаносомы птиц в мозгу. При сонной б-ни человека наблюдается выход трипаносом из сосудов в вещество головного мозга, в мышцу сердца и размножение их в спинномозговой жидкости. Особое положение занимает Schizo-trypanum cruzi, образующая лейшманиевидные формы в клетках ретикуло-гистиоцитарной системы всех органов и в мышечных элементах сердца. Картина на вскрытии погибших от трипаносомоза животных обычно следующая исхудание, отеки, кровоизлияния, увеличение селезенки (в особенности в остром периоде) и лимф. желез. В центральной нервной системе обнаруживаются периваскулярные инфильтрации из круглоклеточных и плазматических элементов, в нервных стволах отеки, дегенерация и мелкоклеточная инфильтрация. Иммунологические взаимоотношения между макро – и микроорганизмами при трипаносомо-зах в значительной степени напоминают отношения, имеющие место при спирохетозах. После заражения наступает первый период размножения возбудителя. К определенному сроку развивается некоторая степень иммунитета, сопровождающаяся появлением в крови специфических антител; наступает массовая гибель паразитов. Часть трипаносом однако выживает, но оказывается измененной в своей антигенной структуре и нечувствительной к антителам первого порядка. Эти серорезистентные рецидивные трипаносомы, размножаясь, ведут к образованию новых антител. Снова гибнет большинство трипаносом, а затем снова появляются трипаносомы с измененными антигенными свойствами. Такая сложная игра продолжается в течение всей инфекции. Антигенные свойства трипаносом меняются не только при образовании рецидивных штаммов в одном и том же животном, но, как показал Кроо (Кгоб), и при перевивке с одного вида животного на другой. Противотрипаносомные антитела обладают ли-тическим и аглютинирующим действием. Эти свойства могут быть обнаружены in vitro. Начальный стадий действия трипанолизинов может быть уловлен феноменом нагрузки бляшек Биццоцеро (Рикенберг). При смешении крови, содержащей трипаносомы, с гомологичной трипанолитической сывороткой происходит адсорпцдя трипапосомами антител, влекущая за собой изменение физ.-хим. свойств перипласта. Поверхность трипаносом при этом становится как бы липкой, а так как трипаносомы на пек-рое время еще сохраняют, свою подвижность, то они легко приходят в соприкосновение с плавающими в крови кровяными бляшками и слипаются С’ этими элементами, к-рые легко пристают к клеточным элементам с поврежденным поверхностным слоем (ср. роль бляшек крови в образовании тромба). В указанных условиях трипаносомы легко облепляются и другими корпускулярными элементами и легко прилипают и к эритроцитам и к лейкоцитам (феномен atlachement). На позднейших стадиях инфекции могут обнаруживаться антитела, не обладающие непосредственным трипаноцидным действием, но задерживающие их развитие (Taliaferro). Особый интерес представляют трипаноцид-ные свойства нормальной сыворот